Mitophagy is the destruction of mitochondria within a cell by specialised ‘garbage disposal’ units called lysosomes. ‘Selective mitophagy’ is the destruction of malfunctioning, defective or simply aged mitochondria. Selective mitophagy is a hot topic today. As long ago as 1978 in my postgraduate days we demonstrated that in senescent rats a significant subpopulation of mitochondria were present in aged livers which were not found when similar aged livers were caused to regenerate following partial hepatectomy1.
Since then it has become accepted from multiple studies that old, damaged, usually enlarged mitochondria inhabit senescent cells and are responsible in signalling the multivarious chain reaction of events that precipitate cell death. This process called apoptosis is now regarded as a partial blessing in old age as a clearing out of the redundant burdensome poorly performing cells but I am more interested in the problem of weeding out the old, poor functioning mitochondria leaving the ‘younger’ to proliferate and predominate? This is, to all intents and purposes, nothing less than rejuvenation … the holy grail of gerontology.
The selection of mitochondria for their ‘fitness’ is not unknown. Small mammals such as rats and mice have typically short life spans ( 2-3yrs in captivity) and we are accustomed to thinking that size matters with regard to mammal longevity: mice, dogs, cats, apes, humans, elephants are roughly in order of mass and lifespan. But how we should envy the Pigeon; it has a body mass not greater than a rat but a life span of over twenty years... and don’t get me started on Parrots who hold the record at 92 yrs. Pigeons, like most birds have very good mitochondria compared to our own. They produce fewer reactive oxidative species (R.O.S) and basically are just more efficiently coupled2.
However there are flying mammals too; furry flying mice-sized creatures known as bats. You guessed it, they have long life spans ( >10yrs) and yes they have good mitochondria too. They are not born with these bird-like mitochondria though. Before they fly theirs are much as they would be in any small mammal but after they fly it’s a different story. It’s hard not to conclude that there was a selection process for the best going on here3.
We are not likely to be able to subject our systems to the energetic stress of flying in order to weed out the less efficient mitochondria, it would kill us, but maybe there are other ways?
Recent fascinating work on the urolithins (related to the tannins and other phenolics in red wines) from pomegranate seeds I think provides not only some clues but also possible mechanism for selective mitophagy. Urolithin A has been shown to extend the life of the simple nematodes and the mechanism is believed to be the result of selective mitophagy4. But how?
How do you label a mitochondrion for destruction? Old, degenerating mitochondria have many ways of signalling their approaching end but these apoptotic signals mean the end of the cell too.
We need to label the mitochondria for destruction without tripping the death chain reactions.
In a previous blog ‘Mitochondrial morphology and Ageing.’5, I put forward a long cherished point of view that enlarged senescent mitochondria were that way in order to maintain their threshold membrane potential against a background of reduced e-/H+ throughput, and increased leaks across the membrane. I used an electronics model of the mitochondrion as capacitor with voltage, current, capacitance and leakage as normal parameters. Basically if you reduce surface area (capacitance) you can maintain the threshold voltage (membrane potential) in the face of lower throughput ( current) and back flow due to dielectric leakage.
In other words the old mitochondria are working but don’t push them as they are close to the edge.
With this in mind look at the urolithin molecule from pomegranates:

It’s a poly-phenol as is the molecule below:
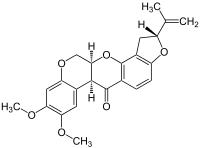
This is rotenone, a potent inhibitor of mitochondria,
which prevents electron flow past Complex I.
It is in many ways ( see bottom left rings) very similar to urolithin.
Or may be you prefer your molecules simpler?
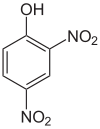
This is DNP (di-nitro phenol) a potent de-coupler of mitochondria.
Both of these molecules are severe toxins. Rotenone is used as a pesticide and scarily and sometimes fatally DNP can be used by those seeking to lose fat weight!
Rotenone (in my electronics analogue) would decrease the current flow through the capacitor and DNP would increase the leak to ground. In either case a mitochondrial capacitor struggling to maintain its threshold membrane potential would be badly compromised.
Maybe then the signal for mitophagy is the blinking on and off of the threshold membrane potential, a kind of ‘drowning not waving’ signal to the lysosomes that these mitochondria are not up to the job. This kind of mechanism clearly would work well as the young bat took to flying and the less fit mitochondria failed to keep up.
Unlike in the bat where ‘stress’ is extreme energetic demand, urolithin mediated selection is being achieved by a hormetic mechanism. A ‘little poison does you good’?
I think that this is the most promising of avenues to explore with regard to rejuvenating mitochondrial populations and thereby rejuvenating the organism. Humans cannot regenerate organs as can an old rat its liver, we cannot (when old) survive extreme energetic stress therefore hormesis seems like a good bet. But what is the best hormesis for mitochondrial rejuvenation… now there’s a question.
- PhD Thesis: Biochemistry of Ageing in Rat Liver Mitochondria: p 165-170:awarded July 1980: online Jan 2016
- Oxygen, The Molecule that made the World: Nick Lane 2002 Oxford University Press pp255-257
Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents, Nature Medicine 22,879–888, (2016)
Ann N Y Acad Sci. 2004 Jun;1019:506-8.Testing the free radical theory of aging in bats. Brunet Rossinni AK
- Mitochondrial morphology and Ageing: January 2015 Spannermans Edublog www.spannerman2.co.uk
